W organizmie człowieka serotonina (5-hydroksytryptamina, 5-HT) pełni rolę zarówno neuroprzekaźnika, jak i hormonu. Ze względu na budowę chemiczną należy do grupy amin biogennych. Wykazuje działanie plejotropowe w ośrodkowym układzie nerwowym (OUN) i poza nim dzięki istnieniu zróżnicowanych pod względem budowy, rozmieszczenia i mechanizmów transmisji sygnału receptorów serotoninowych. Serotonina uwolniona do przestrzeni międzysynaptycznej może oddziaływać na autoreceptory presynaptyczne bądź na heteroreceptory występujące na komórkach postsynaptycznych. Jej działanie zostaje zahamowane poprzez ponowny wychwyt do zakończeń nerwowych. Wśród funkcji jakie pełni, wyróżnia się regulację: pracy serca, krążenia krwi, stanów emocjonalnych, zachowań seksualnych, snu, pobierania pokarmu, a także funkcji poznawczych i uczucia bólu.
1. Historia
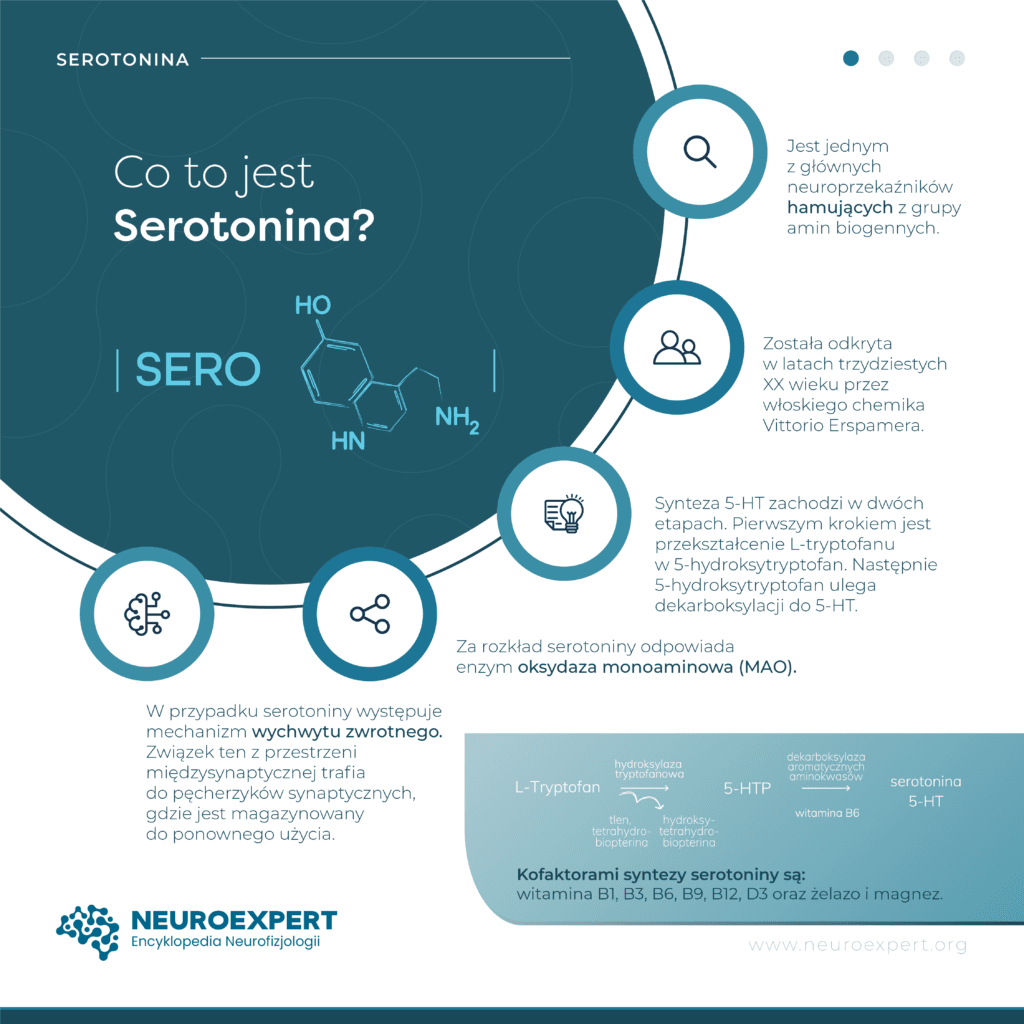
Serotonina została odkryta w latach trzydziestych XX wieku przez włoskiego chemika Vittorio Erspamera. Naukowiec dokonał izolacji związku o właściwościach kurczących miocyty z ziarnistości komórek chromochłonnych błony śluzowej jelita, nadając mu nazwę enteroaminy. Kilka lat później amerykańscy badacze: fizjolog Irvine Page i biochemik Maurice Rapport, prowadząc niezależne badania, również skutecznie zidentyfikowali substancję stymulującą skurcz mięśni gładkich naczyń krwionośnych. W wyniku prowadzenia dalszych badań w 1952 roku stwierdzono, że wszyscy oni odkryli tę samą substancję. Od tego czasu oba opisane związki funkcjonują pod wspólną nazwą: serotonina. W języku angielskim pojęcie “serotonine” funkcjonuje jako akronim. Składają się na niego dwa słowa: surowica (ang. serum), ponieważ substancja występuje w surowicy krwi, oraz przymiotnika wazotoniczny (ang. vasotonic) odnoszącego się do napięcia naczyń krwionośnych.
2. Synteza serotoniny
Szacuje się, że niemal 90% serotoniny w organizmie człowieka jest produkowane w jelitach. Reakcje syntezy odbywają się z udziałem mikrobioty jelitowej oraz specyficznych enzymów obecnych w komórkach enterochromafinowych błony śluzowej jelita. Tak wyprodukowana serotonina nie przekracza bariery krew-mózg, dlatego w ośrodkowym układzie nerwowym jedyne źródło serotoniny stanowią neurony serotoninergiczne. Są one rozproszone w całym OUN, jednak występują szczególnie obficie w korze mózgowej, hipokampie, ciele migdałowatym, warstwie podstawnej zwojów bocznych, pokrywie wzrokowej, jądrach: kolankowatym bocznym oraz nadskrzyżowaniowym, a także w rogach przednich i istocie szarej części grzbietowej rdzenia kręgowego. Od ciał tych komórek odchodzą aksony, które to z kolei rozgałęziają się w specyficzny dla nich sposób. Aksony obejmują swoim zasięgiem istotę szarą znajdującą się w mózgu oraz w rdzeniu kręgowym.
Prekursorem serotoniny jest L-tryptofan (kwas 2-amino-3-indolopropanowy), aminokwas aromatyczny, który nie jest syntetyzowany przez organizm człowieka i musi być przyjmowany wraz z posiłkiem. Dlatego jego biodostępność jest czynnikiem ograniczającym syntezę serotoniny. Tryptofan przenika przez barierę krew-mózg poprzez aktywny system transportu aminokwasów. Dlatego cząsteczki tryptofanu muszą konkurować o miejsce wiązania z innymi substratami: leucyną, lizyną i metioniną.
Synteza 5-HT zachodzi w dwóch etapach. Pierwszym krokiem jest przekształcenie L-tryptofanu w 5-hydroksytryptofan (ang. 5-hydroxytryptophan, 5-HTP) w obecności cząsteczki tlenu. Reakcja ta przebiega przy udziale enzymu hydroksylazy tryptofanu (ang. tryptophan hydroxylase, TPH) określanej również mianem monooksygenazy L-tryptofanu (ang. L-tryptophan-5-monooxygenase). Kofaktorami niezbędnymi do właściwego funkcjonowania tego enzymu są: tetrahydrobiopteryna (ang. tetrahydrobiopterin, BH4), będąca donorem protonów, i magnez. Dodatkowo do prawidłowego metabolizmu BH4 ważne są kwas foliowy (witamina B9) i witamina C, umożliwiające jego regenerację.
Podczas drugiego etapu syntezy serotoniny 5-hydroksytryptofan ulega dekarboksylacji do 5-HT. Reakcja ta zachodzi pod wpływem enzymu dekarboksylazy 5-hydroksytryptofanu (ang. 5-hydroxytryptophan decarboxylase) nazywanego inaczej dekarboksylazą L-aminokwasów aromatycznych (ang. aromatic L-amino acid decarboxylase, AADC). Dla prawidłowego działania tego enzymu niezbędna jest aktywna forma witaminy B6, fosforan pirydoksalu (ang. pyridoxal phosphate, PLP).
3. Receptory serotoniny
Receptory serotoniny stanowią zróżnicowaną grupę obejmującą transportery, kanały jonowe bramkowane ligandem, ale największa grupa receptorów serotoninowych należy do nadrodziny receptorów sprzężonych z białkiem G. Obecnie zidentyfikowano siedem podtypów receptorów 5-HT (od 5-HT1 do 5-HT7) na podstawie wyników badań klonowania. Dalszy podział uzyskano na podstawie ich właściwości strukturalnych i farmakologicznych.
3.1. Receptory 5-HT1
Receptory 5-HT-1 należą do grupy sprzężonej z białkami G. Ulegają ekspresji w obrębie ciała komórki i dendrytów neuronów serotoninergicznych. W obrębie rodziny receptorów 5-HT-1 wyróżnia się pięć podtypów: A, B, D, E, F. Receptory 5-HT1A i 5-HT1D pełnią funkcję autoreceptorów. Oznacza to, że charakteryzują się zdolnością do hamowania transdukcji sygnału na zasadzie ujemnego sprzężenia zwrotnego. Może się to odbywać na dwa sposoby: poprzez hiperpolaryzację błony presynaptycznej w wyniku otwarcia kanałów potasowych i zamknięcia kanałów wapniowych lub poprzez wychwyt zwrotny serotoniny. Znanym agonistą receptora 5-HT1A jest buspiron, zaś antagonistą johimbina.
Podtyp B receptora 5-HT1 wykazuje ekspresję szczególnie w obszarach mózgu takich jak: zwoje podstawy mózgu, prążkowie i kora czołowa. Z kolei receptory 5-HT1E występują głównie w skorupie ogoniastej oraz mniej licznie w ciele migdałowatym, korze czołowej i gałce bladej. Wysoką koncentrację receptora 5-HT1F obserwuje się w szwie grzbietowym, hipokampie i korze mózgu. Jest on także obecny w prążkowiu, wzgórzu i podwzgórzu. Pobudzenie receptorów 5-HT1B, 5-HT1E, 5-HTF skutkuje działaniem hamującym na błonę presynaptyczną w mechanizmie sprzężenia zwrotnego ujemnego i hamowania aktywności cyklazy adenylowej. Do antagonistów receptora 5-HT1B należy johimbina.
3.2. Receptory 5-HT2
Receptory 5-HT2 są sprzężone z białkami G, których enzymem docelowym jest fosfolipaza C. Pośredniczą one w efektach pobudzających poprzez zmniejszenie przewodnictwa dla jonów potasu. Podrodzina receptora 5-HT2 składa się z trzech podtypów: 5-HT2A, 5-HT2B i 5-HT2C. W obrębie OUN receptory 5-HT2 znajdują się w niektórych obszarach przodomózgowia, takich jak: warstwy czwarta i piąta kory nowej, kora gruszkowata, a także guzek węchowy.
3.3. Receptory 5-HT3 i 5-HT4
Receptory 5-HT3 obejmują grupę kanałów sodowo-potasowych bramkowanych ligandem. Cecha ta wyróżnia je od innych typów receptorów serotoninowych, które są sprzężone z białkami G. Receptory 5-HT3 ulegają ekspresji m.in. w korze śródwęchowej, ciele migdałowatym, polu najdalszym pnia mózgu i jądrze pasma samotnego rdzenia przedłużonego. Aktywacja receptorów 5-HT3 w OUN powoduje wzrost przepuszczalności błony dla jonów sodowych i potasowych. Sprawia to, że membrana jest nieprzepuszczalna dla kationów dwuwartościowych, co prowadzi do depolaryzacji neuronu.
Transmisja sygnału przez receptory 5-HT4 odbywa się natomiast z udziałem wtórnego przekaźnika jakim jest cykliczny adenozynomonofosforan (cAMP). Znanymi agonistami tych receptorów są metoklopramid i cisapryd.
3.4. Receptory 5-HT5, 5-HT6 i 5-HT7
Receptory 5-HT5 i 5-HT7 są sprzężone z białkiem G, których enzymem docelowym jest cyklaza adenylowa. Wśród receptorów 5-HT5 wyróżnia się dwa podtypy: 5-HT5A i 5HT5B. Ekspresja tego podtypu receptora zachodzi głównie na komórkach glejowych. Jednak stwierdza się je także w korze mózgowej, opuszce węchowej, hipokampie, móżdżku i warstwie istoty białej zwanej uzdeczką.
Receptory 5-HT6 wykryto w korze mózgowej, hipokampie, prążkowiu, ciele migdałowatym, jądrze półleżącym i guzku węchowym. Z kolei receptory 5-HT7 występują w warstwach 1–3 kory mózgowej, w neuronach przegrody hipokampa, wzgórza, podwzgórza i wzgórka górnego, a także w środkowo-przyśrodkowej części ciała migdałowatego i istocie szarej okołowodociągowej.
4. Usuwanie serotoniny z synapsy
W trakcie fali depolaryzacji cząsteczki serotoniny uwolnione do szczeliny synaptycznej wiążą się m.in. z receptorami sprzężonymi z białkami G, obecnymi na błonie presynaptycznej lub postsynaptycznej neuronów. Aktywacja autoreceptorów presynaptycznych hamuje dalsze uwalnianie serotoniny, podczas gdy pobudzenie receptorów postsynaptycznych, za pośrednictwem przekaźników wtórnych, stymuluje wychwyt zwrotny serotoniny. 5-HT trafia z powrotem do komórki przy udziale transportera serotoniny (ang. serotonin transporter, SER-T). SER-T wykorzystuje jako siłę napędową gradient elektrochemiczny jonów sodowych i potasowych. Znanymi inhibitorami SER-T są leki z grupy selektywnych inhibitorów zwrotnego wychwytu serotoniny (ang. selective serotonin reuptake inhibitor, SSRI). Są one stosowane m.in. w leczeniu depresji i mogą poprawiać nastrój właśnie poprzez zwiększenie dostępności serotoniny w synapsach.
We wnętrzu neuronu serotonina jest transportowana do pęcherzyków synaptycznych, gdzie jest magazynowana do ponownego użycia. Proces ten jest regulowany przez białko znane jako transporter pęcherzyków synaptycznych (ang. vesicular monoamine transporter, VMAT), którego aktywność jest zależna od obecności jonów magnezu. VMAT hamowany jest przez rezerpinę, lek stosowany przez wiele lat w leczeniu nadciśnienia tętniczego. Po zakończeniu transmisji sygnału, serotonina może ulec katabolizmowi w cytozolu neuronu za pośrednictwem specyficznych enzymów.
5. Katabolizm serotoniny
Za rozkład serotoniny odpowiada enzym oksydaza monoaminowa (ang. monoamine oxidase, MAO). Jej izoenzymy MAO-A i MAO-B obecne w OUN i innych tkankach ciała. 5-HT, która została wytworzona w jelitach, trafia do krążenia wrotnego, a następnie jest katabolizowana w wątrobie i płucach. Główny szlak przemian katabolicznych serotoniny inicjowany jest przez izoenzym A. MAO przekształca 5-hydroksytryptaminę do aldehydu 5-hydroksyindolooctowego (ang. 5-hydroxyindole acetaldehyde). Kofaktorem w tej reakcji jest związany kowalencyjnie z cząsteczką MAO dinukleotyd flawinoadeninowy (ang. flavin adenine dinucleotide, FAD), pełniący rolę przenośnika protonów i elektronów. W kolejnym etapie 5-hydroksyindolooctan jest utleniany z pomocą dehydrogenazy aldehydowej do kwasu 5-hydroksy-3-indolooctowego (ang. 5-hydroxyindoleacetic acid, 5-HIAA), będącego głównym metabolitem serotoniny. Powstały związek jest wydalany z organizmu wraz z moczem i może być wykorzystywany jako biomarker w laboratoryjnych badaniach diagnostycznych nowotworów czynnych hormonalnie np. rakowiaka.
Katabolizm serotoniny zachodzi także w wyniku reakcji metylacji i dezaminacji oksydacyjnej, prowadzonych przez odpowiednie enzymy: metylotransferazę serotoniny oraz oksydazę aminową. Ścieżka tych przemian prowadzi do powstania 5-metoksyindolooctanu (ang. 5-methoxyindole acetate, 5-MIAA). W ośrodkowym układzie nerwowym serotonina może też być przekształcana do melatoniny w szyszynce.
6. Znaczenie serotoniny
Potocznie serotonina nazywana jest „hormonem szczęścia”, co oznacza, że w dużej mierze przyczynia się do regulacji nastroju. Swoim działaniem obejmuje przede wszystkim ośrodkowy układ nerwowy oraz układ pokarmowy. Z powodu występowania w tym ostatnim odpowiada za apetyt. Za jej pośrednictwem regulowana jest także temperatura ciała oraz ciśnienie krwi. Bardzo ważne jest, że neuroprzekaźnik ten, wraz z melatoniną, reguluje sen, czyli dba o dobrą regenerację organizmu.
Najbardziej niepożądanym skutkiem obniżenia się poziomu serotoniny w ogranizmie jest pojawienie się depresji. Spadek powoduje także bezsenność, co w konsekwencji przejawia się brakiem energii, ospałością a także może być przyczyną agresji. Osoba z niedoborem serotoniny nie potrafi myśleć racjonalnie. Do negatywnych skutków może doprowadzić także nieprawidłowy metabolizm tego neuroprzekaźnika. W tym przypadku pojawiają się zaburzenia behawioralne. Do zaburzeń tych zaliczyć można: zaburzenia obsesyjno-kompulsywne, stany lękowe, a w niektórych przypadkach nawet bulimię.
7. Działanie serotoniny w OUN
7.1. Sen i czuwanie
Serotonina wywiera swój wpływ na sen poprzez oddziaływanie na neurony obecne w jądrach nadskrzyżowaniowych, które są uważane za część wewnętrznego zegara biologicznego. Szczególną rolę w tych procesach pełnią neurony jąder szwu w obrębie których produkowana jest serotonina. Te komórki nerwowe są najbardziej aktywne w fazie czuwania, ich aktywność spada natomiast w czasie snu NREM i zostaje całkowicie zahamowana w fazie REM.
7.2. Procesy poznawcze i emocjonalne
Potocznie serotonina nazywana jest „hormonem szczęścia”, gdyż w dużej mierze przyczynia się do regulacji nastroju. Jeśli chodzi o działanie przeciwlękowe, to odbywa się ono głównie poprzez aktywację receptorów 5-HT1A i 5-HT2C. Serotonina reguluje zachowania seksualne oraz odczuwanie bólu za pośrednictwem projekcji neuronów odpowiednio: z jąder dziobowych i jąder ogonowych układu serotoninergicznego. Ponadto 5-HT nadzoruje procesy poznawcze poprzez obecne w strukturach limbicznych receptory 5-HT7.
7.3. Kontrola apetytu
Serotonina reguluje apetyt i wpływa na układ nagrody poprzez receptor 5-HT2C. Zapobiega napadom nadmiernego głodu oraz chroni przed wybieraniem niewłaściwych pokarmów. Gdy aktywność 5-HT w mózgu jest zrównoważona, nie dochodzi do przejadania się. Serotonina może również pośredniczyć w wyzwalaniu odruchu wymiotnego poprzez pobudzanie receptorów 5-HT3 zlokalizowanych w obrębie pola najdalszego pnia mózgu oraz w aferentnych włóknach nerwu błędnego przewodu pokarmowego. Znanym antagonistą tych receptorów jest ondansetron, który stosowany jest jako lek o działaniu przeciwwymiotnym.
7.4. Regulacja wydzielania innych substancji
Neurony serotoninergiczne wpływają na uwalnianie hormonów przysadki mózgowej, zwłaszcza prolaktyny i gonadotropiny. Receptory HT1A i 5-HT2 są zaangażowane w regulację wydzielania czynników uwalniających kortykotropinę, uczestnicząc w regulacji osi podwzgórze-przysadka-nadnercza (ang. hypothalamic–pituitary–adrenocortical axis, HPA). Serotonina zwiększa wydzielanie płynu mózgowo-rdzeniowego, aktywując receptory 5-HT2C w splotach naczyniówkowych.
8. Obwodowe efekty działania serotoniny
8.1. Układ sercowo-naczyniowy
Serotonina odgrywa rolę w regulacji ciśnienia krwi, oddziałując na receptory 5-HT1 i 5-HT2 w obrębie mięśni naczyń krwionośnych. W zależności od rodzaju receptorów obecnych w mięśniach gładkich, serotonina może prowadzić do zwężenia lub rozszerzenia łożyska naczyniowego. Aktywacja receptorów 5-HT1B i 5-HT1D rozszerza tętnice mózgowe. Te podtypy receptorów są celem leków takich jak tryptany stosowane w leczeniu migreny. Pobudzając receptory 5-HT2 na powierzchni śródbłonka naczyniowego, serotonina wywołuje skurcz mięśni gładkich i zwężenie łożyska naczyniowego. Antagonista receptorów 5-HT2, ketanseryna, wykorzystywany jest w leczeniu nadciśnienia tętniczego.
W odpowiedzi na niedotlenienie organizmu serotonina może nasilać przekazywanie sygnałów przez mitogenny receptor 5-HT2B na komórkach śródbłonka tętnic płucnych. Zwiększa to proliferację komórek mięśni gładkich naczyń, a tym samym przyczynia się do rozwoju nadciśnienia płucnego. Podobne działanie zaobserwowano u pacjentów, stosujących lek fenfluraminę w terapii otyłości i depresji. W tym przypadku 5-HT może stymulować rozwój nadciśnienia płucnego w mechanizmie wzrostu aktywności transportera serotoniny.
Serotonina obecna w krwiobiegu uczestniczy w procesach hemostazy, aktywując receptory 5-HT2 w obrębie płytek krwi. Trombocyty mogą również wychwytywać 5-HT z osocza za pośrednictwem transportera serotoniny, magazynować ją w ziarnistościach i uwalniać razie potrzeby. Działanie tego hormonu na układ krążenia obejmuje także wzrost siły i częstości skurczów serca poprzez zwiększenie wewnątrzkomórkowego stężenia wapnia w kardiomiocytach.
8.2. Przewód pokarmowy
W przewodzie pokarmowym serotonina aktywuje receptory 5-HT4, co skutkuje m.in. przyspieszeniem opróżniania żołądka, przyspieszeniem pasażu jelitowego oraz zwiększonym wydzielaniem soków trawiennych. Ponadto 5-HT reguluje aktywność wydzielniczą trzustki, stymuluje wychwyt glukozy w tkance mięśniowej oraz lipogenezę w tkance tłuszczowej i wątrobie.
8.3. Układ mięśniowo-szkieletowy
Serotonina reguluje napięcie mięśni nie tylko w obrębie naczyń krwionośnych, ale i również innych narządów, w tym oka. Przez skurcz mięśni ciała rzęskowego 5-HT rozszerza źrenice, co z kolei prowadzi do wzrostu ciśnienia wewnątrzgałkowego.
Receptory 5-HT obecne są w każdym rodzaju komórek budujących kości: osteoblastach, osteocytach i osteoklastach. Wyniki badań pokazują, że serotonina aktywuje transbłonową lipoproteinę LRP5 (ang. LDL receptor-related protein 5), która pełni kluczową rolę w procesach proliferacji, różnicowania, adhezji i migracji osteoblastów. Neuroprzekaźnik ten generuje również transdukcję sygnału szeregu białkowego Wnt. Prawidłowe funkcjonowanie tego białka jest podstawą profilaktyki osteoporozy.
9. Zaburzenia
Dysfunkcje układu serotoninergicznego są przyczyną powstawania procesów patologicznych, które z kolei są fundamentalną podstawą chorób psychicznych, m.in. depresji, schizofrenii, zaburzeń łaknienia, zaburzeń snu, a także stanów lękowych.
Najbardziej niepożądanym skutkiem obniżenia się poziomu serotoniny w organizmie jest pojawienie się depresji. Niedobór serotoniny pojawiający się u osób cierpiących z powodu depresji prowadzi do hipersomnii, czyli uczucia senności pomimo prawidłowo przespanej nocy. Spadek poziomu serotoniny powoduje także bezsenność, co w konsekwencji przejawia się brakiem energii, ospałością, a także może być przyczyną agresji. Osoba z niedoborem serotoniny nie potrafi myśleć racjonalnie.
Do negatywnych skutków może doprowadzić także nieprawidłowy metabolizm tego neuroprzekaźnika. W tym przypadku pojawiają się zaburzenia behawioralne. Do zaburzeń tych zaliczyć można: zaburzenia obsesyjno-kompulsywne, stany lękowe, a w niektórych przypadkach nawet bulimię. W kontekście zaburzeń łaknienia niedostateczny poziom serotoniny prowadzi do nadmiernego wydzielania hormonu koncentrującego melaninę (ang. melanin-concentrating hormone, MCH) w podwzgórzu. Skutkuje to nadmiernym odczuwaniem głodu i wzmożoną potrzebą spożycia pokarmu najczęściej pod postacią niezdrowych przekąsek (bogatych w węglowodany i tłuszcze), których spożycie prowadzi do wzrostu insuliny. Jej nadmiar sprzyja z kolei magazynowaniu tłuszczu w organizmie, a więc i powstawaniu otyłości.
10. Przyczyny zaburzeń
10.1. Przyczyny niedoboru serotoniny:
- niski poziom witaminy B6 w organizmie – aktywna forma tej witaminy, czyli fosforan-5-pirydoksalu odgrywa kluczową rolę w przemianie L-tryptofanu w serotoninę;
- dieta uboga w produkty zawierające L-tryptofan – aminokwas ten jest zwiazkiem prekursorowym w syntezie serotoniny. Nie jest on jednak produkowany w organizmie, dlatego musi być do niego dostarczany wraz z pożywieniem;
- “konkurencja aminokwasowa” – L-tryptofan jest stosunkowo małym aminokwasem, dlatego przy spożywaniu wysokobiałkowych produktów, pierwszeństwo w przekraczaniu bariery krew-mózg mają większe cząsteczki aminokwasów. Aby konkurencję tę wyeliminować, dobrze jest L-tryptofan przyjmować w towarzystwie węglowodanów;
- nadmiar teaniny – jest to aminokwas obecny w liściach herbaty. Posiada on zdolność do przekraczania bariery krew-mózg oraz hamowania neuronów wydzielających serotoninę w ośrodkowym układzie nerwowym;
10.2. Przyczyny nadmiaru serotoniny:
- rakowiak – jest to nowotwór neuroendokrynny, przyjmujący postać hormonalnie czynną. Nazwą tą określa się guzy powstające w organizmie, których komórki pobierają znaczną ilość L-tryptofanu i zamieniają go w serotoninę. Produktem rozkładu tego neurotransmitera jest kwas 5-hydroksyindolooctowy wydalany z moczem. Wzrost tego metabolitu w moczu jest więc markerem pozwalającym na diagnozę nowotworu;
- długotrwałe stosowanie SSRI (selektywne inhibitory wychwytu zwrotnego serotoniny, ang. Selective Serotonin Reuptake Inhibitors) lub IMAO (inhibitory monoaminooksydazy)
10.2.1. Zespół serotoninowy:
Przyjęcie zbyt dużej dawki substancji (leku lub suplementu diety) podnoszącej poziom serotoniny w ośrodkowym układzie nerwowym może skutkować rozwojem zespołu serotoninowego, który może prowadzić nawet do śmierci (do 1999 roku stwierdzono 23 przypadki śmiertelne). Objawy zespołu serotoninowego to:
- ból lub zawroty głowy
- halucynacje
- sztywność i skurcze mięśni
- uczucie niepokoju i zagubienia
- podwyższenie temperatury ciała
- nadmierne pocenie się
- przyspieszenie czynności serca
- nadciśnienie tętnicze
- bezsenność
- drgawki
- rozszerzenie źrenic
- nudności i wymioty
- biegunka
11. Wpływ suplementów na aktywność serotoniny
| SUPLEMENT | Synteza i uwalnianie | Oddziaływanie na receptory | Usuwanie z synapsy | Uwagi |
| 5-HTP (Griffonia simplicifolia) | + | prekursor | ||
| Brahmi (Bacopa monnieri) | + | stymulacja syntezy | ||
| Kurkumina (Curcuma longa) | + | inhibitor MAO | ||
| Dziurawiec (Hypericum perforatum) | + | + | zwiększenie stężenia receptorów, inhibitor wychwytu zwrotnego, inhibitor MAO | |
| Eleuthero (Eleutherococcus senticosus) | + | inhibitor MAO | ||
| Garcinia (Garcinia cambogia) | + | zwiększenie stężenia prekursora w mózgu | ||
| Goji (Lycium barbarum) | + | źródło kofaktora do syntezy | ||
| Kofeina | + | zwiększenie stężenia receptorów | ||
| L-teanina (Camellia sinensis) | +/- | stymulacja syntezy / w nadmiarze – obniża poziom serotoniny | ||
| Piperyna (Piper nigrum) | + | inhibitor MAO | ||
| Różeniec (Rhodiola rosea) | + | inhibitor MAO | ||
| SAM-e | modulacja aktywności | modulacja aktywności | modulacja aktywności | dokładny mechanizm działania nieznany |
| Tarczyca bajkalska (Scutellaria baicalensis) | + | inhibitor MAO | ||
| Winpocetyna | + | stymulacja uwalniania |
(+) – zwiększenie aktywności serotoniny
(-) – zmniejszenie aktywności serotoniny
12. Podsumowanie