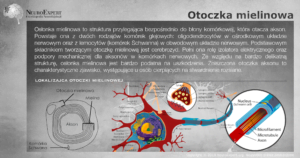
Osłonka mielinowa, nazywana inaczej osłonką rdzenną, to struktura otaczająca włókna nerwowe. Pełni ona kluczową rolę w przewodzeniu impulsów nerwowych oraz oraz stanowi podporę mechaniczną dla aksonów komórek nerwowych. Włókna nerwowe zawierające otoczkę mielinową nazywamy rdzennymi, a te które jej nie zawierają bezrdzennymi.
1. Historia
W roku 1854 Rudolf Virchow skonstruował termin „mielina” na podstawie greckiego słowa oznaczającego „szpik” (myelos), w celu opisania struktury występującej obficie w mózgu. Virchow postulował, że mielina jest wytwarzana przez neurony i pełni rolę izolującej masy. Sto lat później badania histologiczne przeprowadzone przez Pio Del Rio Hortegę i Wildera Penfielda sugerowały, że to oligodendrocyty odpowiadają za produkcję mieliny. Dzięki zastosowaniu barwienia węglanem srebra byli oni w stanie zidentyfikować oligodendrocyty z ich cienkimi wypustkami połączonymi z osłonkami mielinowymi. Stwierdzono, że oligodendrocyty są obecne w znacznym stopniu w istocie białej, lecz występują również w istocie szarej, przeważnie w obrębie pęczków nerwowych, czasami także w okolicach naczyń krwionośnych.
2. Budowa
Osłonka mielinowa otacza niektóre włókna nerwowe, tworząc spiralne warstwy wokół aksonów. Stanowi ona przedłużenie błony komórkowej oligodendrocytów w ośrodkowym układzie nerwowym (OUN) i komórek Schwanna w obwodowym układzie nerwowym.
Osłonka rdzenna składa się w około 80% z lipidów oraz w 20% z białek. Wśród tłuszczów dominują cholesterol, cerebrozydy, głównie galaktozyloceramid oraz fosfolipidy takie jak lecytyna (fosfatydylocholina). Do białek wchodzących w skład osłonki mielinowej zalicza się białko mieliny podstawowej (ang. myelin basic protein, MBP) i białko proteolipidowe (ang. proteolipid protein, PLP), które stabilizują warstwy lipidowe i utrzymują integralność osłonki.
Skład osłonki mielinowej może się różnić w zależności od lokalizacji w układzie nerwowym. W OUN obserwuje się większą zawartość glikolipidów i mniejszą ilość fosfolipidów w osłonkach mielinowych niż w obwodowym układzie nerwowym. Struktura mieliny zależy od składu lipidów. Wysoki poziom wielonienasyconych kwasów tłuszczowych (ang. polyunsaturated fatty acids, PUFA) w osłonkach w obrębie obwodowego układu nerwowego jest związany z luźną strukturą mieliny. Z kolei wysokie stężenie cholesterolu jest wymagane do tworzenia zwartej struktury mieliny.
Prędkość przewodzenia sygnału nerwowego jest bezpośrednio związana ze średnicą aksonu, dlatego aksony cechujące się grubą i gęstą osłonką mielinową przewodzą impulsy szybko. Takie aksony tworzą włókna w obrębie łuków odruchowych. Natomiast neurony z cienkimi wypustkami, otoczone luźną osłonką mielinową, przewodzą sygnały wolniej i występują głównie w autonomicznym układzie nerwowym
Wyniki badań wskazują, że skład lipidów budujących osłonki mielinowe może ulegać zmianom. To odkrycie otwiera nowe możliwości dla tworzenia innowacyjnych strategii terapeutycznych w obszarze leczenia chorób neurodegeneracyjnych, poprzez modulację zarówno składu ilościowego, jak i jakościowego substancji budulcowych mieliny.
3. Funkcje
3.1. Przyspieszenie przewodzenia impulsów
Osłonka mielinowa umożliwia znaczne przyspieszenie przewodzenia impulsów nerwowych, pozwalając im osiągnąć prędkości sięgające nawet 100 m/s (we włóknach bezmielinowych prędkość ta wynosi 0,1 m/s). Na całej długości osłonki otaczającej akson w odstępie co ok. 1 mm pojawia się przewężenie o długości ok. 1 μm, nazywane węzłem Renviera. W tym obszarze osłonka nie występuje. Przewodzenie impulsów między przewężeniami odbywa się w sposób skokowy, co znacząco przyspiesza propagację impulsów nerwowych. Ponadto w obrębie węzłów Ranviera występują kanały sodowe i potasowe zależne od napięcia, biorące udział w depolaryzacji i inicjacji potencjału czynnościowego.
Przewodzenie impulsów nerwowych wiąże się z wydatkiem energetycznym. Dzięki obecności osłonki mielinowej komórki nerwowe oszczędzają energię. Impulsy nie muszą być bowiem generowane na całej długości aksonu, lecz tylko w obszarach pomiędzy węzłami Ranviera. W ten sposób impuls elektryczny przemieszcza się skokowo wzdłuż aksonu, przechodząc od jednego przewężenia do kolejnego. Dzięki temu dany odcinek jest pokonywany znacznie szybciej niż w przypadku braku osłonki mielinowej, przy jednoczesnym zachowaniu pełnej siły impulsu.
3.2. Ochrona i izolacja
Układ warstw lipidowych nadaje osłonce mielinowej jej izolacyjne właściwości. Dzięki temu chroni ona aksony przed uszkodzeniami mechanicznymi i chemicznymi, zapobiega dyfuzji jonów przez błonę komórkową oraz minimalizuje ewentualne zakłócenia elektryczne między aksonami skupionymi w pęczki. Umożliwia to bardziej skuteczną i dokładną transmisję sygnałów nerwowych.
4. Powstawanie
Mielinizacja, czyli proces tworzenia osłonki mielinowej, stanowi złożony i precyzyjnie skoordynowany mechanizm, zachodzący na różnych etapach rozwoju komórek nerwowych. Uważa się, że w OUN mielinizacja obejmuje włókna o średnicy ok. 0,3 μm, a w obwodowym do 1 μm. Aksony o średnicy 0,2 µm lub mniejszej zazwyczaj nie posiadają mieliny. Jeden z możliwych czynników wyjaśniających związek między mielinizacją a średnicą aksonów to niezdolność mieliny do wyginania się poza pewną krzywiznę, co utrudnia owijanie aksonów małego kalibru.
4.1. Inicjacja
Wpływ czynników wzrostu oraz sygnałów regulacyjnych inicjuje aktywację genów związanych z procesem mielinizacji w komórkach prekursorowych. To prowadzi do przekształcenia ich w oligodendrocyty lub komórki Schwanna, które są odpowiedzialne za tworzenie osłonek mielinowych w OUN i obwodowym układzie nerwowym. W tym procesie istotną rolę pełni sieć transkrypcyjna, którą kontroluje czynnik transkrypcyjny PPARγ.
4.2. Mechanizm
Mielinizacja jest procesem dynamicznym, którego przebieg różni się w zależności od wieku i potrzeb organizmu. Rozpoczyna się około piątego miesiąca życia płodowego w korzeniach ruchowych rdzenia kręgowego. Następnie obejmuje regiony kontrolujące złożone funkcje w obrębie płatów czołowych mózgu. W większości osłonka mielinowa powstaje w ciągu pierwszych dwóch lat życia. Uważa się, że w istocie białej mózgu proces ten może trwać dłużej nawet przez 2-3 dekady.
Podczas mielinizacji błona komórkowa oligodendrocytów oraz komórek Schwanna owija się wokół aksonów, tworząc osłonkę mielinową. Z reguły pojedynczy oligodendrocyt mielinizuje wiele sąsiednich aksonów, podczas gdy komórka Schwanna – tylko jeden akson. Proces ten zachodzi w sposób nieciągły, co oznacza, że pomiędzy dwiema kolejnymi komórkami Schwanna lub oligodendrocytami w równych odstępach występują miejsca pozbawione osłonki mielinowej.
Mielinizacja jest regulowana przez białka odpowiedzialne za adhezję oraz kształtowanie osłonki. Ponadto istotną rolę w tym procesie mają jony wapnia, wpływające na dynamikę cytoszkieletu, który bierze udział w owijaniu osłonki wokół aksonów.
5. Lokalizacja a różnice w mielinizacji
Pomiędzy różnymi obszarami układu nerwowego obserwuje się znaczące zróżnicowanie w czasie i tempie mielinizacji, np. aksony w nerwie wzrokowym przechodzą proces mielinizacji w dynamicznym tempie i we wczesnym etapie rozwoju. Z kolei mielinizacja aksonów w korze czołowej postępuje wolniej i zachodzi w późniejszym okresie.
Aksony w rdzeniu kręgowym i nerwie wzrokowym są mielinizowane jednoetapowo, co stwarza sprzyjające warunki do osiągnięcia maksymalnej prędkości przewodzenia impulsów. Z kolei aksony w korze mózgowej przechodzą mielinizację w dwóch fazach. Pierwsza z nich polega na nieregularnym rozmieszczeniu segmentów mieliny wzdłuż włókien nerwowych. Drugi etap, prawdopodobnie wywołany przez bodźce sensoryczne, uzupełnia powstałe luki, co zapewnia odpowiednią komunikację między neuronami a oligodendrocytami.
6. Czynniki wpływające na proces mielinizacji
Proces mielinizacji włókien nerwowych jest ściśle regulowany przez wiele czynników, do których należą cytokiny, neurotransmitery i inne molekuły sygnałowe. Wpływają one na różne etapy mielinizacji, co w konsekwencji determinuje efektywność i tempo tego procesu. Niemniej jednak mechanizmy molekularne kontrolujące ten proces pozostają wciąż słabo poznane.
Wśród kluczowych czynników promujących mielinizację wymienia się białko oddziałujące ze Smad1 (ang. Smad Interacting Protein, Sip1), znane również jako Zeb2 oraz czynniki transkrypcyjne oligodendrocytów (ang oligodendrocyte transcription factor, Olig). Mogą one wzajemnie na siebie oddziaływać. Nadekspresja Olig1 i Olig2 pobudza ekspresję genu białka Sip1, będącego kluczowym regulatorem przejścia prekursorów oligodendrocytów do dojrzałych komórek, które wytwarzają osłonkę mielinową w OUN. Jednocześnie Sip1 jest represorem transkrypcji genów grupy białek morfogenetycznych kości (ang. bone morphogenetic protein, BMP), które są uważane za czynniki hamujące mielinizację. Szlak sygnałowy z udziałem BMP a także szlak Wnt/β-katenina blokują różnicowanie prekursorów oligodendrocytów. Mogą one również utrudniać regenerację i remielinizację włókien nerwowych.
Wyniki badań sugerują, że aktywność elektryczna neuronów może stymulować mielinizację poprzez przekazywanie sygnałów, które promują rozwój i funkcjonowanie osłonki mielinowej. W niektórych obszarach układu nerwowego, np. w mózgu, proces mielinizacji może być modyfikowany przez doświadczenie i naukę. Intensywna aktywność neuronów w odpowiedzi na różne bodźce może wpływać na tempo mielinizacji i organizację osłonek mielinowych.
7. Patologia
Zaburzenie organizacji struktury osłonki mielinowej lub jej uszkodzenie określa się mianem demielinizacji. Te zmiany mogą być konsekwencją procesów starzenia, urazów, działania toksycznych substancji, czynników infekcyjnych i genetycznych, czy też chorób neurodegeneracyjnych i autoimmunologicznych.
Uszkodzenia integralności osłonki mielinowej w ośrodkowym i obwodowym układzie nerwowym są również obecne w schorzeniach związanych z zaburzeniami metabolizmu lipidów. Akumulacja lipidów lub ich prekursorów może prowadzić do lipotoksyczności i uszkodzenia osłonki mielinowej, a w konsekwencji do różnych schorzeń układu nerwowego. Defekty tego typu mogą objawiać się zmniejszoną produkcją osłonki mielinowej (hipomielinizacja), nieprawidłową strukturą (dysmielinizacja) lub jej brakiem.
Degeneracja osłonki mielinowej aksonów jest obserwowana w przypadku stwardnienia rozsianego, które jest chorobą zapalną o podłożu autoimmunologicznym. W przebiegu tego schorzenia, obserwuje się degradację oligodendrocytów, obecność blaszek zapalnych i obszarów pozbawionych mieliny oraz nacieki wokół naczyń krwionośnych składające się z makrofagów i limfocytów. Zmiany te skutkują uszkodzeniem i utratą aksonów. W wyniku tego prędkość przewodzenia impulsów nerwowych może spadać nawet dziesięciokrotnie, a w skrajnych przypadkach może dojść do całkowitego przerwania przekazywania sygnałów nerwowych.
Uszkodzenie osłonek mielinowych obserwuje się także w przebiegu neurotoksyczności związanej z lekami przeciwnowotworowymi, znanej również jako „chemobrain”. Wysokie stężenia 5-fluorouracylu, powszechnie stosowanego środka chemioterapeutycznego, okazują się toksyczne zarówno dla komórek progenitorowych OUN, jak i dla oligodendrocytów, zarówno w badaniach in vitro, jak i in vivo. Przyczyny tego schorzenia obejmują takie mechanizmy jak stres oksydacyjny i stan zapalny, obniżony poziom neuroprzekaźników, zmniejszoną neurogenezę oraz zaburzenie produkcji osłonki rdzennej.
8. Podsumowanie